









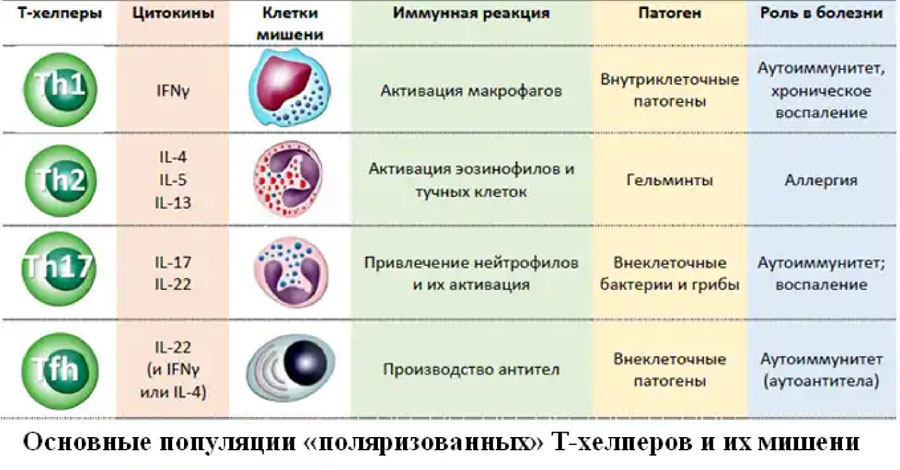
 Среди всех клеток систем врожденного и приобретенного иммунитета Т-хелперы (Тh) выделяются особенным разнообразием выполняемых функций, что находит свое отражение в крайне высокой гетерогенности данной популяции (табл.23). Исходно на основании продукции цитокинов среди CD3+CD4+ лимфоцитов выделяли клетки, способные к синтезу IFNγ и получившие название Т-хелперов 1 типа (Тh1), и клетки, названные Т-хелперами 2 типа (Тh2) и синтезировавшие IL-4. Далее были описаны Th17 и фолликулярные Т-хелперы (Tfh). Мы остановимся только на этих четырех типах клеток, так как их реальное существование не подвергается сомнению. Следует отметить, что с завидной регулярностью появляются работы, свидетельствующие о возможности перехода Th из одной популяции в другую в зависимости от микроокружения, типа получаемых цитокиновых сигналов метаболизма клеток и широчайшего спектра других факторов.
Среди всех клеток систем врожденного и приобретенного иммунитета Т-хелперы (Тh) выделяются особенным разнообразием выполняемых функций, что находит свое отражение в крайне высокой гетерогенности данной популяции (табл.23). Исходно на основании продукции цитокинов среди CD3+CD4+ лимфоцитов выделяли клетки, способные к синтезу IFNγ и получившие название Т-хелперов 1 типа (Тh1), и клетки, названные Т-хелперами 2 типа (Тh2) и синтезировавшие IL-4. Далее были описаны Th17 и фолликулярные Т-хелперы (Tfh). Мы остановимся только на этих четырех типах клеток, так как их реальное существование не подвергается сомнению. Следует отметить, что с завидной регулярностью появляются работы, свидетельствующие о возможности перехода Th из одной популяции в другую в зависимости от микроокружения, типа получаемых цитокиновых сигналов метаболизма клеток и широчайшего спектра других факторов.
Т-Хелперы 1 типа (Th1)
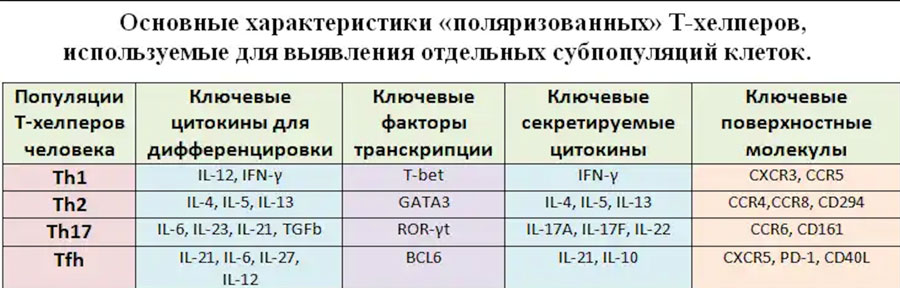 Для Th1 характерна продукция провоспалительных цитокинов IFNγ и TNFα, а также IL-2 и IL-15, наличие транскрипционного фактора T-bet и экспрессия на поверхностной мембране CXCR3, CCR5 и рецептора для IL-18 (IL-18R). Ключевую роль в «поляризации» Th0 в Th1 играет IL-12, синтезируемый в лимфоидной ткани активированными дендритными клетками, хотя IL-18 и IL-33 в присутствии IL-12 также способствуют появлению CD3+CD4+ клеток со свойствами Th1. Основной задачей Th1 является активация тканевых макрофагом и мигрировавших в периферические ткани моноцитов с целью повышения эффективности уничтожение поглощенных этими клетками патогенов. Основным же факторов, обеспечивающим реализацию этих функций, является IFNγ. К важнейшим свойствам данного цитокина относятся:
Для Th1 характерна продукция провоспалительных цитокинов IFNγ и TNFα, а также IL-2 и IL-15, наличие транскрипционного фактора T-bet и экспрессия на поверхностной мембране CXCR3, CCR5 и рецептора для IL-18 (IL-18R). Ключевую роль в «поляризации» Th0 в Th1 играет IL-12, синтезируемый в лимфоидной ткани активированными дендритными клетками, хотя IL-18 и IL-33 в присутствии IL-12 также способствуют появлению CD3+CD4+ клеток со свойствами Th1. Основной задачей Th1 является активация тканевых макрофагом и мигрировавших в периферические ткани моноцитов с целью повышения эффективности уничтожение поглощенных этими клетками патогенов. Основным же факторов, обеспечивающим реализацию этих функций, является IFNγ. К важнейшим свойствам данного цитокина относятся:
- IFNγ является ключевым цитокином, вызывающим активацию макрофагов;
- IFNγ стимулирует микробицидную активность макрофагов и их поляризацию в сторону М1;
- IFNγ способствует переключению класса синтезируемых В-лимфоцитов антител с IgM на некоторые изотипы IgG (например, у мышей – на IgG2a или IgG2c), подавляя синтез и секрецию IgE;
- IFNγ способствует «поляризации» «наивных» Тх в сторону Тх1, подавляя Тх2 и Тх17 программы дифференцировки;
- IFNγ способствует увеличению интенсивности презентации антигенов через МНС II, тем самым, усиливая клеточный ответ.
При гиперфункции Th1, связанной с продукцией цитокинов (в первую очередь, IFNγ и TNFα), в организме могут формироваться очаги хронического воспаления, вызывающие повреждения окружающих тканей (как это показано при формировании гранулем). Более того, формирование клонов аутореактивных Th1 связано с развитием органно-специфических и системных аутоиммунных заболеваний.
Т-Хелперы 2 типа (Th2)
Th2 синтезируют IL-4, IL-5, IL-6, IL-9, IL-10 и IL-13, отвечающие за регуляцию клеточного и гуморального иммунного ответа, в их цитоплазме и ядре обнаруживается транскрипционный фактор Gata3, а на своей мембране они несут CD294, CCR3, CCR4, CCR8 и некоторые дополнительный антигены. При отсутствии IL-12 в окружающей ткани, но наличии IL-4 и IL-33, Th0 дифференцируются в Th2 клетки. Основной задачей Th2 является активация тканевых тучных клеток и мигрировавших в периферические ткани из крови базофилов и эозинофилов с целью повышения эффективности уничтожение крупных многоклеточных патогенов (в первую очередь, гельминтов) и грибов. Поэтому, помимо активации лейкоцитов цитокины Th2 влияют на функционирование глакомышечных клеток трубчатых органов, эндотелий сосудов и тканевые макрофаги. Основные эффекты цитокинов Th2:
- IL-4 стимулирует переключение синтезируемых антител В-клетками с IgM на IgE;
- IL-4 играет ведущую роль в формировании Th2 из «наивных» Тh и является важнейшим аутокринным ростовым факторов для дифференцировки Тh2;
- IL-4, совместно с IL-13, отвечает за формирование и функциональную активность М2 макрофагов в очаге воспаления;
- IL-4 (и IL-13) стимулирует перистальтику кишечника, а сам IL-13 повышает секрецию слизи эпителиальными клетками ВДП и ЖКТ;
- IL-4 и IL-13 привлечение лейкоцитов (в первую очередь, эозинофилов) в очаг воспаления за счет активации эндотелия сосудов микроциркуляторного русла и секреции хемокинов.
- IL-5, совместно с IL-3 и GM-CSF, действует как фактор, стимулирующий эозинофилы, причем, IL-5 стимулирует пролиферацию и дифференцировку предшественников эозинофилов, тогда как IL-3 и GM-CSF действуют на более ранних стадиях развития. IL-5 также стимулирует дифференцировку В-клеток в плазматические клетки, секретирующие IgA.
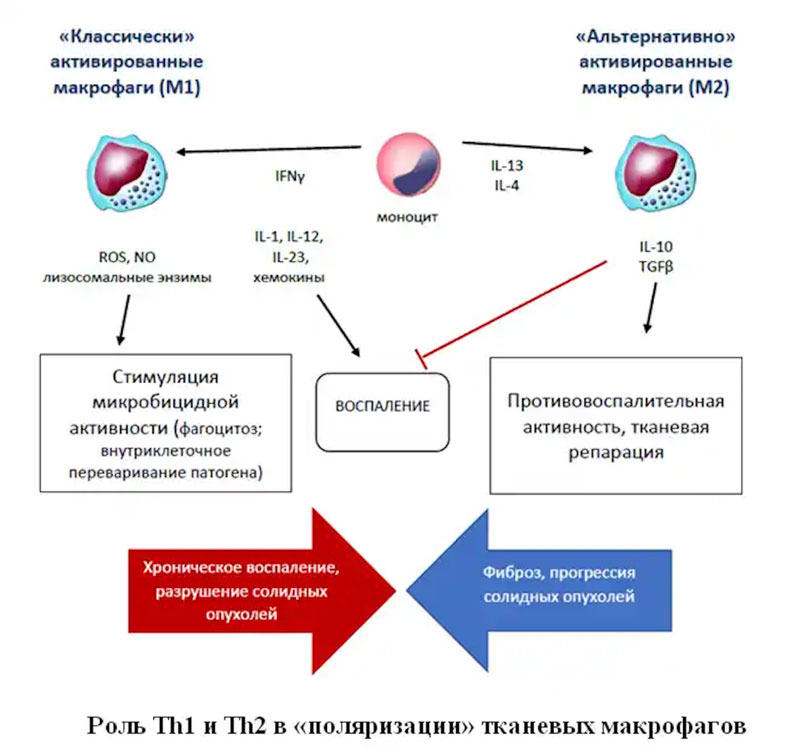 Гиперактивация Th2 и продукция ими цитокинов для активации тучных клеток и базофилов тесто связана с развитием аллергических заболеваний, что находит широкое клиническое применение – например, блокада IL-5 при помощи терапевтических антител при бронхиальной астме). Вместе с тем, именно Th2 стимулируют «поляризацию» Тканевых макрофагов в сторону М2, что способствует запуску процессов репарации ткани при гельминтозах, хотя гиперпродукция цитокинов может сопровождаться формированием очагов фиброза в тканях.
Гиперактивация Th2 и продукция ими цитокинов для активации тучных клеток и базофилов тесто связана с развитием аллергических заболеваний, что находит широкое клиническое применение – например, блокада IL-5 при помощи терапевтических антител при бронхиальной астме). Вместе с тем, именно Th2 стимулируют «поляризацию» Тканевых макрофагов в сторону М2, что способствует запуску процессов репарации ткани при гельминтозах, хотя гиперпродукция цитокинов может сопровождаться формированием очагов фиброза в тканях.
Т-Хелперы 3 типа (Th17)
Th17 были впервые описаны в 2005 как клетки с фенотипом CD3+CD4+, способные к синтезу и секреции IL-17A, IL-17F, IL-21 и IL-22. Формирование Th17 из «наивных» клеток у мыши и человека происходит под действием таких цитокинов как TGF-β, IL-1β, IL-6, IL-21 и IL-23. На на поверхностной мембране Th17 обнаружена высокая плотность таких антигенов как CXCR4, CCR6 и CD161. Основной задачей Th17 является активация тканевых и мигрировавших в периферические ткани из крови нейтрофилов с целью повышения эффективности уничтожение внеклеточных патогенов за счет фагоцитоза. Поэтому, помимо активации лейкоцитов цитокины Th17 влияют на функционирование клеток эпителиальных пластов с целью повышения из барьерных функций, а также на активацию клеток эндотелия сосудов, тканевых макрофагов и фибробластов. К основным эффектам цитокинов Th17 относятся:
- IL-17 активирует макрофаги и дендритные клетки на продукцию провоспалительных медиаторов, что способствует индукции воспаления;
- Действуя на клетки эпителиальных пластов IL-17 (совместно с IL-22) усиливает секрецию ими слизи и антимикробных пептидов, а также хемокинов, что увеличивает защитные свойства барьерных тканей;
- IL-17 стимулирует фибробласты соединительной ткани на продукцию провоспалительных цитокинов и хемокинов, что способствует увеличению эффективности клеточных реакций в очаге проникновения патогена;
- Совместно с другими цитокинами IL-17 обладает рядом системных эффектов, связанных с активацией клеток микроциркуляторного русла, гепатоцитов печени, мышечных клеток сердца и скелетной мускулатуры.
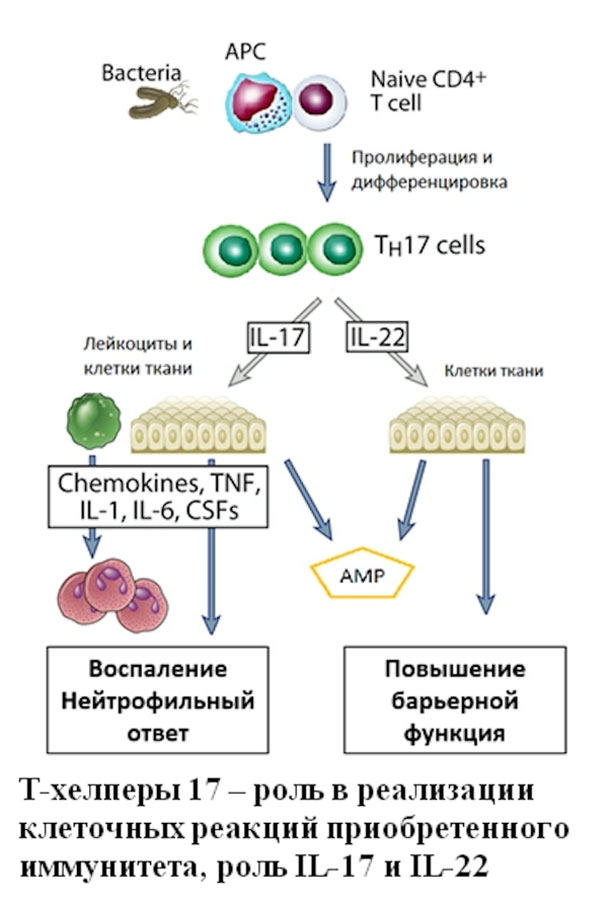 В целом, Тh17 играют ведущую роль в защите от внеклеточных патогенов — бактерий и грибов – вызывая активацию нейтрофилов («нейтрофильное» воспаление) и клеток барьерных тканей. Более того, Тh17 совместно с Тh1 регулируют функциональную активность фагоцитов (нейтрофилов и моноцитов) при реализации клеточно-опосредованных реакций врожденного иммунитета. Однако, гиперактивация Тh17 может служить причинной развития широкого спектра патологических состояний, в основе которых находятся неконтролируемые воспалительные процессы в различных тканях — псориаз, воспалительное заболевание кишечника, ревматоидный артрит и рассеянный склероз. Именно поэтому Th17 являются мишенью для различных биологических препаратов, направленных на поляризующие и эффекторные цитокины (анти-IL-6, анти-IL-17, анти-IL-23, анти-TNF терапевтические моноклональные антитела).
В целом, Тh17 играют ведущую роль в защите от внеклеточных патогенов — бактерий и грибов – вызывая активацию нейтрофилов («нейтрофильное» воспаление) и клеток барьерных тканей. Более того, Тh17 совместно с Тh1 регулируют функциональную активность фагоцитов (нейтрофилов и моноцитов) при реализации клеточно-опосредованных реакций врожденного иммунитета. Однако, гиперактивация Тh17 может служить причинной развития широкого спектра патологических состояний, в основе которых находятся неконтролируемые воспалительные процессы в различных тканях — псориаз, воспалительное заболевание кишечника, ревматоидный артрит и рассеянный склероз. Именно поэтому Th17 являются мишенью для различных биологических препаратов, направленных на поляризующие и эффекторные цитокины (анти-IL-6, анти-IL-17, анти-IL-23, анти-TNF терапевтические моноклональные антитела).
По результатам взаимодействия между АПК и «наивным» Т-лимфоцитов, а также клональной экспансии и дифференцировки, формируется, как уже отмечалось выше несколько типов Т-хелперов, которые различаются по своим свойствам:
- Т-лимфоциты центральной памяти:
– способны пролиферировать и быстро дифференцироваться (обычно 3-5 дней) в эффекторные клетки;
– локализуются в периферических лимфоидных органах, а, благодаря способности к «рециркуляции», регулярно посещаю («патрулируют») периферические лимфоидные органы различной локализации, живут годами – отвечают за долговременный иммунитет;
– нуждаются в презентации антигена в периферических лимфоидных органах и ко-стимуляции для проявления своих свойств и клональной экспансии;
– не обладают выраженными эффекторными свойствами.
- Т-лимфоциты эффекторной памяти:
– постоянно находятся в периферических тканях – потенциальных участках повторного проникновения патогена;
– быстро активируются и увеличивают уровень продукции цитокинов и других эффекторных молекул;
– не нуждаются в презентации антигена в периферических лимфоидных органах и ко-стимуляции;
– обладают низкой способностью к пролиферации.
- Эффекторные Т-лимфоциты:
– время жизни 10-20 дней;
– локализуются преимущественно в периферических тканях;
– не способны к пролиферации.
– обладают выраженными эффекторными свойства, играют ведущую роль в продукции цитокинов, необходимых для эффективного функционирования эффекторных клеток (моноцитов и макрофагов в случае Th1; тучных клеток, базофилов и эозинофилов в случае Th2; клеток соединительной ткани, эпителиальных платов и нейтрофилов в случае Th17) врожденного иммунитета.








